Neue Moleküle und Wege der Antigenpräsentation bei der Tuberkulose
Forschungsbericht (importiert) 2003 - Max-Planck-Institut für Infektionsbiologie
Neue Moleküle und Wege der Antigenpräsentation bei der Tuberkulose
Tuberkulose stellt auch heute noch eine der bedrohlichsten Infektionskrankheiten weltweit dar. Ein Drittel der Weltbevölkerung ist mit dem bakteriellen Erreger Mycobacterium tuberculosis infiziert. Dies führt pro Jahr in 8-9 Millionen Fällen zur klinischen Manifestation der Tuberkulose, typischerweise in der Lunge, von denen etwa 2 Millionen tödlich enden. Diese Daten verdeutlichen eindringlich die Bedeutung der Tuberkulose für das Gesundheitswesen und den Bedarf für einen wirkungsvollen Impfstoff.
Meist führt die Infektion mit dem Erreger jedoch nicht zur Erkrankung und der Infektionsverlauf wird durch das körpereigene Immunsystem wirkungsvoll kontrolliert. Diese immunologische Kontrolle bewirkt aber nur die "Einkapselung" der Erreger in Granulomen; die sterile Elimination der Keime gelingt nicht. Die Infektion bleibt daher lebenslang bestehen und kann auch nach Jahren eine Erkrankung auslösen.
Im Zentrum der Infektabwehr stehen Fresszellen des angeborenen Abwehrsystems, die Makrophagen, die zwar einerseits durch Mykobakterien infiziert werden, aber andererseits durch das Zytokin Interferon-g aktivierbar sind und dann intrazelluläre Bakterien abtöten oder deren Wachstum hemmen. Die zweite Achse der Abwehr wird von T-Lymphozyten gebildet, die zum erworbenen Immunsystem gehören und spezifisch mykobakterielle Antigene erkennen, die ihnen von "professionellen" antigenpräsentierenden Zellen (wie dendritischen Zellen (DZ) und Makrophagen) angeboten werden. Dabei spielen sog. CD4-T-Lymphozyten die entscheidende Rolle bei der T-Zell-Antwort, obwohl auch andere T-Zell-Populationen wesentlich zur Immunabwehr gegen Tuberkulose beitragen. Unsere hier vorgestellen Arbeiten konzentrieren sich auf CD8-T-Zellen, die wie CD4-T-Zellen Proteinantigene erkennen, sowie auf CD1-restringierte T-Lymphozyten, die antigenspezifisch auf Lipide reagieren.
Das Problem der CD8-T-Zell-Aktivierung bei der Tuberkulose
Obwohl CD8-T-Lymphozyten in erster Linie für die Virusabwehr zuständig sind, werden sie für die Abwehr bestimmter Bakterien, so auch M. tuberculosis, dringend benötigt. Mykobakterien-spezifische CD8-T-Zellen können aus dem Gewebe infizierter Patienten isoliert werden und Experimentaltiere, denen CD8 T-Zellen fehlen, zeigen einen deutlich verringerten Schutz gegen Tuberkulose.
CD8-T-Zellen differenzieren nach Antigenerkennung und folgender Aktivierung zu zytotoxischen T-Lymphozyten (ZTL), die infizierte Zielzellen durch sezernierte Proteine wie Perforin lysieren können. Dadurch werden intrazelluläre Bakterien freigesetzt und für direkte Attacken durch ZTL angreifbar. Weiterhin produzieren ZTL Interferon-g, das wiederum infizierte Phagozyten zur potenten Keimabtötung mittels reaktiver Sauerstoff- und Stickstoffmetabolite aktiviert. Die antigenspezifische Aktivierung der CD8-T-Zellen gab lange Zeit Rätsel auf. Unsere Untersuchungen zeigten, dass Mykobakterien nach Eintritt in den infizierten Makrophagen in einem abgeschlossenen Kompartiment, dem frühen Phagosom, verbleiben und daher zur Aktivierung von CD8-T-Zellen ungeeignet sind. Experimente mit markierten Erregern ergaben, dass das Mykobakterien-Phagosom nahezu hermetisch gegenüber dem Zytoplasma abgeriegelt ist. Da aber Antigene, die von CD8-T-Zellen erkannt werden, normalerweise im Zytoplasma vorliegen müssen, warf dies die Frage auf, wie die mykobakteriellen Antigene an ihren Bestimmungsort gelangen, um CD8-T-Zellen zu stimulieren. Das Phänomen, dass Antigene von der Zelle, in der sie primär anfallen, nicht präsentiert werden, sondern erst abgegeben und von einer zweiten Zelle, meist einer DZ, aufgenommen und präsentiert werden müssen, ist als "Cross-priming" bekannt. Aufzuklären, ob und ggf. über welchen Mechanismus "Cross-priming" bei Tuberkulose stattfindet, war das nächste Ziel unserer Experimente.
Apoptotische Vesikel transportieren mykobakterielle Antigene zu DZ
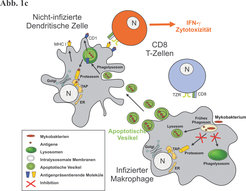
Unsere In-vitro-Versuche zeigten, dass die Infektion mit M. tuberculosis in Makrophagen einen programmierten Zelltod, die "Apoptose" auslöst. Dabei handelt es sich um den geordneten Prozess des Sterbens einzelner Zellen, der physiologisch in jedem Organismus die Zellzahl, trotz gleichzeitiger Neubildung von Zellen, konstant hält. Die herrschende Meinung ging bisher von einem nachteiligen Effekt der infektionsinduzierten Apoptose aus, da mit dem Makrophagen ein Hauptakteur der Abwehr beseitigt wird. Wir konnten zeigen, dass die infizierte Zelle während der Apoptose Vesikel abschnürt (Abb. 1a), die von umliegenden DZ aufgenommen werden. Genauere Untersuchungen der apoptotischen Vesikel (Abb. 1b) ergaben, dass sie zahlreiche dominante mykobakterielle Antigene wie z.B. das 19kDa-Lipoprotein und das sog. Antigen85 enthalten. Die von den DZ aufgenommenen apoptotischen Vesikel aktivierten mykobakterien-reaktive CD8-T-Zell. Dabei stand eine lysosomale Prozessierung der Vesikel in den DZ im Vordergrund, da die Behandlung mit Lysosomen-Inhibitoren wie Bafilomycin die T-Zell-Aktivierung unterband, wohingegen Proteasomen-Inhibitoren einen zu vernachlässigenden Effekt auf die T-Zell-Aktivierung hatten. Wurde die Apoptose in den primär infizierten Makrophagen durch Zugabe eines globalen Kaspase-Inhibitors gehemmt, so wurden der Antigentransfer und die CD8-T-Zell-Aktivierung unterbunden. Da die mykobakteriellen Antigene von der Sackgasse im Phagosom des infizierten Makrophagen, über die apoptotischen Vesikel hin zur DZ mit erfolgreicher Antigenpräsentation und T-Zell-Aktivierung einen Umweg nehmen müssen, um überhaupt CD8-T-Zellen stimulieren zu können, bezeichnen wir diesen neuen Weg der Antigenpräsentation als "Detour pathway" (Abb. 1c).


Detour pathway in vivo
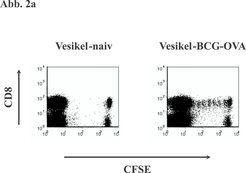
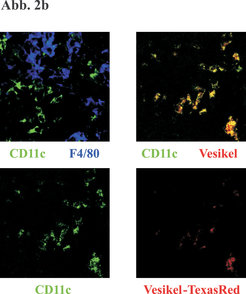
Um zu untersuchen, ob "Cross-priming" über apoptotische Vesikel nicht nur ein lokales Geschehen der Antigenverteilung darstellt, wie es im peripheren entzündeten Gewebe, in dem gehäuft Zellen sterben, stattfindet, sondern tatsächlich einen genuin immunologischen Prozess repräsentiert, der das Immunsystem miteinschließt, dehnten wir unsere Experimente auf ein In-vivo-Modell aus. Dabei transferierten wir eine definierte Population von CD8-T-Zellen in Empfängermäuse und immunisierten diese Tiere mit apoptotischen Vesikeln von mykobakterien-infizierten Makrophagen. Bei den CD8-T-Zellen handelte es sich um Lymphozyten mit Spezifität für Ovalbumin (OVA). Wir infizierten Makrophagen für die Vesikelpräparation mit rekombinanten Mykobakterien, die OVA als Surrogat-Antigen exprimieren. In diesem definierten System ließ sich nachweisen, dass apoptotische Vesikel von infizierten Zellen nach subkutaner Immunisierung in der Lage sind, CD8-T-Zellen in den drainierenden Lymphknoten zu aktivieren (Abb. 2a). Nach subkutaner Gabe farbig-markierter Vesikel waren die apoptotischen Vesikel nur in den DZ der drainierenden Lymphknoten wiederzufinden. Wir schließen daraus, dass die DZVesikel in der Peripherie aufnehmen und anschließend in den drainierenden Lymphknoten transportieren (Abb. 2b). Diese Ergebnisse zeigen, dass der "Detour pathway" auch in vivo funktionsfähig ist und das klassische immunologische "Cross-priming" umfasst.


Apoptotische Vesikel und Impfung gegen Tuberkulose
Vakzinierung von Mäusen mit apoptotischen Vesikeln von infizierten Zellen und anschließende Aerosol-Infektion mit M. tuberculosis bewirkte einen Impfschutz vergleichbar dem einer BCG-Immunisierung. Obwohl der BCG-Impfstoff die wichtigsten Tuberkuloseformen nicht verhindern kann, stellt er bis heute den Goldstandard der Vakzinierung gegen Tuberkulose im Tiermodell dar. Damit ist der "Detour pathway" nicht nur biologisch relevant, sondern stellt auch einen weiteren Schritt hin zur rationalen Impfstoffentwicklung dar. Prima vista scheint die Immunogenität der apoptotischen Vesikel dem Konzept gegenläufig, dass apoptotische Vorgänge ohne Entzündung und ohne Aktivierung des Immunsystems ablaufen, da sonst der Organismus permanent auf endogenen Zelltod reagieren müsste und Apoptose damit eng mit Autoimmunität verbunden wäre. Wir müssen bei apoptotischen Vesikeln, die von infizierten Zellen abstammen, jedoch berücksichtigen, dass diese Vesikel per se nicht-entzündlich sind. Andererseits sind sie aber mit einer Vielzahl mykobakterieller Bestandteile ausgerüstet, die nicht nur antigene, sondern auch inflammatorische und adjuvante Eigenschaften vermitteln. Damit werden sie für das Immunsystem als Träger von Fremdmaterialien erkennbar. So konnten wir zeigen, dass apoptotische Vesikel von mykobakterien-infizierten Makrophagen Wirtszellen über die Bindung von Toll-like Rezeptoren (TLR) stimulieren. TLR werden u.a. von DZ exprimiert und dienen der Erkennung konservierter bakterieller Muster. TLR-Bindung induziert Reifung und potente Antigenpräsentation von DZ. Wir sehen diese apoptotischen Vesikel als Träger von Antigenen, antigen-präsentierenden Molekülen und Adjuvanzien, und sprechen ihnen daher eine autonome immunologische Entität zu.
Lipid-spezifische T-Zellen
Die CD1-Familie umfasst ungewöhnliche antigen-präsentierende Moleküle, die von DZ exprimiert werden. Die CD1-Moleküle verfügen über eine antigen-bindende Tasche. Im Gegensatz zur peptidbindenden Grube der wichtigsten antigen-präsentierenden Moleküle der Haupthistokompatibilitätsfamilie dient die CD1-Tasche jedoch der Bindung von Lipiden. Die verschiedenen Mitglieder der CD1-Familie binden unterschiedliche Liganden, u.a. Fettsäuren, Glyzerinester und Sphingolipide. Diese Fette können von CD1-restringierten T-Lymphozyten erkannt werden, die dann als Effektoren ähnliche funktionelle Eigenschaften aufweisen wie konventionelle T-Zellen. Das CD1b-Molekül findet sich primär beim Menschen und bindet vor allem Fette aus der Zellwand von M. tuberculosis wie Mykolsäure, Lipoarabinomannan (LAM) und Glukose-monomykolat (GMM). Anders als beim konventionellen Weg der Antigenpräsentation von Peptiden durch Genprodukte der Haupthistokompatibilitätsfamilie waren die Mitspieler der CD1-Antigenpräsentation bisher unbekannt. Uns gelang die Entdeckung von Saposinen als erste Gruppe von Helfermolekülen bei der Antigenpräsentation über CD1b.
Saposine als fehlendes Bindeglied der Lipid-Präsentation durch CD1
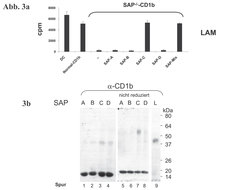
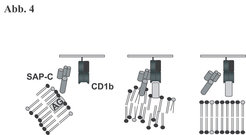
Die wesentliche Frage, die uns zur Suche nach Helfermolekülen bei der Lipidpräsentation trieb, war, wie die biophysikalische Kluft zwischen der hydrophoben Phase der Lipidantigene, die membranorganisiert in der Zelle vorliegen, und der wässrigen Phase, in der die CD1-Proteine exponiert sind, überbrückt werden kann, um die Beladung der CD1-Tasche mit dem Lipid zu gewährleisten. Da bekannt war, dass CD1b und mykobakterielle Antigene wie LAM im Lysosom der Zelle aufeinandertreffen, wählten wir als Kandidaten für unsere Untersuchungen die Sphingolipid Aktivator Proteine (SAP oder Saposine). SAP sind am Abbau und Transport von Fetten im Lysosom beteiligt. Wir untersuchten, ob sie auch bei der Antigenpräsentation eine Rolle spielen. Hierzu transfizierten wir SAP-defiziente Fibroblasten mit CD1b, um die so generierten Zellen als antigenpräsentierende Zellen (APZ) einzusetzen. SAP-defiziente APZ waren nach Inkubation mit Lipidantigen nicht in der Lage, antigen-spezifische, CD1b-restringierte T-Zellen zu stimulieren. Durch Rekonstitution der APZ mit Saposin C und Inkubation mit Lipidantigen wurde die T-Zell-Aktivierung wiederhergestellt (Abb. 3a). Damit war die essentielle Funktion von SAP-C für die Lipid-Präsentation über CD1b belegt. Darüber hinaus konnten wir den dualen Charakter von SAP-C bei der Interaktion mit Lipiden und CD1b aufklären. In Versuchen mit Liposomen erwies sich SAP-C in der Extraktion von Lipidantigen aus Membranen als überaus wirksam. Bindungsstudien mit markierten Saposinen zeigten schießlich, dass SAP-C mit CD1b in der Zelle direkt interagiert (Abb. 3b). Daraus ergibt sich eine Dreiecksbeziehung bei der Lipidantigenpräsentation aus SAP-C, Lipid und CD1b. Unserem Konzept folgend gräbt sich SAP-C in intralysosomale Membranen, destabilisiert deren Membrankomposition, exponiert dabei Lipidantigene und führt sie dem CD1b-Molekül zu, das es gleichzeitig bindet (Abb. 4). Dies zeichnet SAP-C als zentralen Angelpunkt der Lipid-Präsentation und der folgenden Aktivierung CD1b-restringierter T-Lymphozyten aus.